


Najwspanialsza bajka świata
Książka Dawkinsa Najwspanialsze widowisko świata. Świadectwa ewolucji [44] ukazała się 9 lat po książce Wellsa Ikony ewolucji. Byłem przekonany, że Dawkins, skoro pisze o dowodach ewolucji, zaatakuje Wellsa i przedstawi kontrdowody. O dziwo, Dawkins nie wspomniał nawet o tej książce. Nie ma jej też w spisie polecanych lektur.
Dawkins co prawda twierdził, że jeśli prawdziwi naukowcy – tacy jak on – włączą się w dyskusję z kreacjonizmem – będzie to dla nie naukowców dowód, że sprawa kreacjonizmu jest poważna, skoro autorytety naukowe zabierają w niej głos. W związku z tym uważa, że z kreacjonistami nie powinno się nawet rozmawiać. Problem w tym, że Wells nie jest kreacjonistą. Jest naukowcem, który wykazuje, że to co ewolucjoniści uważają za dowody ewolucji wcale nimi nie jest i że ewolucja jest jedynie nieudowodnioną teorią.
Mam wrażenie, że milczenie Dawkinsa wynika raczej z braku kontrargumentów. Żeby coś mówić, trzeba mieć coś do powiedzenia. Dawkins ma rację – jeśli się nie ma nic do powiedzenia należy milczeć.
Czytałem książki Dawkinsa. Zawsze podziwiałem jego swobodny, płynny styl i zdolność do tworzenia skonsolidowanych i pewnie wyglądających teorii. Dawkins doskonale wiedział, że są to tylko słowa, nie poparte żadnymi dowodami, ale za to ładnie brzmiące. Jednym słowem to doskonałe książki z dziedziny science fiction. Dawkins postanowił więc przedstawić – jak sam twierdzi – świadectwa ewolucji, żeby nikt nie mówił, że wszystko to co pisze to jedynie teorie wyssane z palca.
Psy, krowy i kapusta
Dawkins przypomina, że wszelkie zmiany ewolucyjne, zachodzą powoli, stopniowo, dzięki niewielkim mutacyjnym zmianom genetycznym. Dzięki temu istnieją linie ewolucyjne, a np. króliki o krótkich uszach, przekształcają się w króliki o długich uszach i stopniowo coraz bardziej odbiegają od tego królika, którego przyjęliśmy za wzorzec i umieściliśmy na początku naszej linii ewolucyjnej i tak w nieskończoność – w obie strony linii. Idąc w górę – dochodzimy do nowych form ewolucyjnych, idąc w doł do królikopodobnych lub zupełnie niepodobnych do królika przodków naszego królika. Ponieważ zmiany zachodzą w różnych kierunkach ten przodek królików mógł ewoluować w różne inne formy tworząc odgałęzienia ewolucyjne.
Dalej autor podaje przykład, że np. królik i np. leopard (lampart) pochodziły od wspólnego przodka. Nie znaczy to, że króliki wyewoluowały w leopardy albo leopardy wyewoluowały w króliki. Są krewnymi, gdyż pochodzą od wspólnego przodka.
Poniżej przedstawiamy gałąź drzewa ewolucyjnego na której znajduje się zarówno królik jak i leopard. Ponieważ 'królicza’ gałąź i 'leopardzia’ gałąź wyrastają ze wspólnej gałęzi to oba te gatunki musiały mieć wspólnego przodka. Z tej samej gałęzi wyrastają gałęzie lub gałązki naczelnych, prałożyskowców, nietoperzy, pradrapieżnych, drapieżnych , etc.
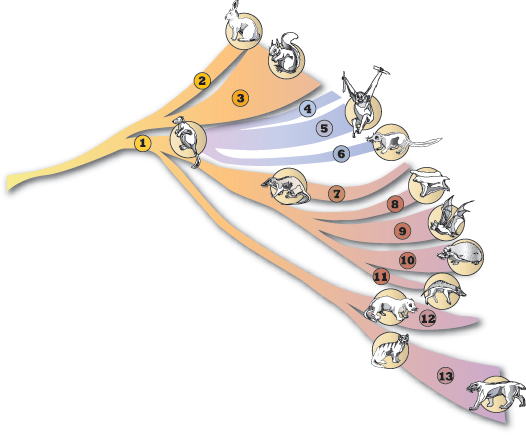
Jeżeli zajęczaki i kotowate miały w ogóle wspólnego przodka, ten przodek musiał w drodze wielu pośrednich stadiów przekształcić się w jednej linii w zajęczaka, w drugiej linii w kotowatego. Pytanie brzmi: jak ten przodek wyglądał. Jeśli był jakoś musiał wyglądać.
Załóżmy cztery możliwości:
- był podobny do współczesnego zająca
- był podobny do współczesnego leoparda
- był podobny do obu naraz – czyli wyglądał jak zającokot czy kotozając mając po połowie cech jednego i drugiego
- był niepodobny do żadnego z nich

Jeśli był podobny do współczesnego zająca lub podobny do współczesnego leoparda to sytuacja była niesymetryczna. Przekształcenie w zająca lub leoparda było łatwe i proste, przekształcenie w drugą z form musiało być dość złożone i wymagało większej liczby kroków. Jeśli był mieszanką cech to przekształcenie w obie formy było jednakowo średnio trudne. Ponieważ leopardy rozmnażają się wolniej niż króliki można przypuścić, że przodek ten był bardzie kotowaty niż zającowaty.
Trzeba podkreślić dwa fakty:
- jeśli ewolucja zachodzi, to formy pośrednie musiały istnieć
- jeśli ewolucja jest powolna i stopniowa – to form pośrednich musiało być bardzo, bardzo dużo, a jeśli było ich bardzo dużo to powinny być łatwo i często znajdowane
Jeżeli przodek zająca i leoparda nie był podobny do żadnego z nich to tym gorzej dla teorii ewolucji, gdyż form pośrednich musiało być jeszcze więcej … Jakkolwiek wyglądał musiał być kręgowcem, czyli posiadał wyodrębnioną głowę, kręgosłup z wyodrębnioną szyją i ogonem, klatkę piersiową i cztery kończyny. Wiemy więc jak w przybliżeniu wyglądał.
Gdybyśmy tego stwora (lub naszego) umieścili na drzewie ewolucyjnym w miejscu pokazanym strzałką moglibyśmy między niego, a końcowe zwierzaki wstawić setki lub tysiące form pośrednich coraz bardziej zbliżających się do swych końcowych postaci. Nie zapominajmy, że ogniwa pokazane tutaj jako końcowe zawierają dziesiątki lub setki gatunków, które nie są pokazane na schemacie
Problem w tym, że w badaniach ewolucyjnych tej gałęzi ewolucyjnej, o której mówimy nigdy i w żadnym przypadku nie znaleziono żadnej formy pośredniej – i to nie tylko na linii zajęczaki – kotowate. Na żadnej z linii w ogóle. Nie znaczy to oczywiście, że nie istniały, ale ich istnienie jest coraz mniej prawdopodobne.
Gdy ewolucjoniści znajdują jakiś kopalny gatunek nadają mu nazwę i umieszczają jako liść na ewolucyjnym drzewie. Dziwne drzewo, które nie ma żadnych gałęzi, gałązek czy pędów – ma wyłącznie liście.
W tym samym rozdziale autor podaje przykłady sztucznej selekcji i hodowli różnych ras psów i udomowionego bydła, nazywając to rzeźbieniem genetycznym.
Nie ma to – oczywiście – wiele wspólnego z ewolucją ani tym bardziej z rzeźbieniem . W procesach hodowlanych zarówno u roślin jak i u zwierząt stosuje się np. chów w pokrewieństwie, czyli rozmnażanie osobników ze sobą spokrewnionych. Jest to brutalna ingerencja w pulę genetyczną gatunku. Prowadzi to do zubożenia puli genowej gatunków, ze względu na homozygotyzację i co z tym związane powstawania nowych korzystnych dla hodowców cech, ale i ujawniania się cech niekorzystnych dla hodowanych zwierząt. Taka selekcja i chów wsobny zachodzi w naturze tylko w ekstremalnych sytuacjach. Do tego co jest w stanie wytworzyć człowiek, natura nigdy nie dopuści.
Szalona podróż do makroewolucji i uroki życia
Autor sporo mówi o udomowieniu wilków, które stały się psami. Po pierwsze nie wiadomo, czy psy i wilki mają w ogóle cokolwiek wspólnego poza względnym podobieństwem. Po drugie sądzę, że ludzie od czasu do czasu znajdowali w lesie samotne wilcze szczenięta – dlatego samotne, bo straciły matkę albo ta matka je porzuciła. Nie miały wyjścia – musiały 'zaufać człowiekowi’. Potem ludzie rozmnażali je w niewoli i nie znały już wilczego życia. A skoro matka ufała człowiekowi, to i jej szczenięta człowiekowi ufały. Hodowały się w pokrewieństwie, gdyż grupy ludzkie były niewielkie i izolowane od siebie, a więc i ich psy (czy wilki) były izolowane od innych. Stąd się wzięły zmiany wilków w psy. Stąd się wzięły zalążki ras. Zmiany genetyczne, które zaszły w tym procesie – są trudno odwracalne. Dlatego watahy zdziczałych psów nie stają się wilkami, ani się z nimi nie zadają. A ponieważ stada się nie łączą, nie ma przepływu genów. To typowy przykład ewolucji wymuszonej przez człowieka podobnie jak wszystkie inne hodowlane odmiany. Różnice między psami a wilkami są głównie behawioralne. Większość psów może dawać potomstwo z wilkami.
Według współczesnych twierdzeń pies nie jest udomowionym wilkiem. Wilk i pies to oddzielne gatunki, które pochodzą od wspólnego przodka – tak więc pies domowy jest udomowionym psem dzikim (który wymarł), a nie udomowionym wilkiem, a więc wszelkie dyskusje na temat udomowienia wilka nie mają racji bytu. Wiadomo, że różne eskimoskie plemiona krzyżowały psy pociągowe z wilkami, aby dodać psom niektórych wilczych cech – przede wszystkim siły i szybkości.
Cisza i powolny czas
Autor pisze o metodach pomiaru czasu używanych w badaniach geologicznych i paleontologicznych.
Metoda słojów drzew zakłada, że istnieje różnica warunków pomiędzy poszczególnymi latami. To zakłada również, że zawsze istniał ten sam klimat, co jest nieprawdopodobne, jeśli mówimy o milionach lat. Współczesne przyrosty drewna mogą nie mieć nic wspólnego z przyrostami drewna w lasach przed setkami milionów lat. Tę metodę można stosować jedynie do 10 tys. lat wstecz.
Metody radioaktywne nie sprawdzają się – jak to podkreśla sam autor – gdyż szczątki zwierzęce występują jedynie w skałach osadowych, a pierwiastki radioaktywne w skałach krystalicznych. Jedyną stosowaną metodą jest metoda węgla C-14. Ta metoda może być stosowana do maksymalnie 60 tys. lat wstecz. Jest nieprecyzyjna ze względu na zmiany zawartości dwutlenku węgla w atmosferze.
Powszechnie przyjmuje się, że skały osadowe leżące na spodzie są – na ogół – starsze niż skały leżące na wierzchu. Skały starsze zawierają szczątki innych gatunków zwierząt, a skały wyżej położone – szczątki innych zwierząt. Np. ssaki nie występują poniżej pewnej warstwy – podobnie jak inne gatunki zwierzęce. Dla ewolucjonistów jest to dowód, że tych zwierząt w niższych warstwach nie było. Dawkins wyśmiewa – nie poparte niczym – twierdzenia kreacjonistów, że osady powstały w czasie potopu, a zoologiczny skład warstw jest związany z szybkością ginięcia poszczególnych rodzajów zwierząt – ginące później znalazły się w wyższych warstwach.
Według geologów procesy fossylizacji trwają setki tysięcy lub nawet miliony lat. Naukowcy ignorują udowodnione fakty znajdowania żywych żab i innych zwierząt w skałach, datowanych według nich na 60 mln lat. Ignorują również udowodnione przykłady fossylizacji osadów zachodzących w okresach miesięcy lub lat.
Czy 6 dni = 14 000 000 000 lat? Czy Pan Bóg w ciągu każdej sekundy dzieła stworzenia tworzył to co według ewolucjonistów natura wytworzyła w ciągu 27000 lat?
Wrócimy jeszcze do tych tematów w dalszym ciągu tej książki.
Masz to przed samym nosem
Autor opisuje spadek długości kłów u ugandyjskich słoni na przestrzeni 35 lat. Pomiary były robione przez myśliwych posiadających uprawnienia do odstrzału. Dawkins skłania się do poglądu, że widzimy 'ewolucję przy pracy’.
To oczywista bzdura. Myśliwy, który dostawał zezwolenie na odstrzał szukał osobnika o jak największych kłach. Ginęło więcej słoni o większych kłach. Zostawały i rozmnażały się słonie o mniejszych rozmiarach kłów. Myśliwy mieli do odstrzału słonie o coraz mniejszych kłach, etc. Jest to typowy przykład sztucznej selekcji dokonywanej w naturalnych warunkach. Selekcję zapewne pogłębiali kłusownicy.
Dawkins zaprzecza tutaj ewolucyjnym poglądom, które mówią, że ewolucja zachodzi w okresach liczonych w setkach tysięcy lat z powodu działania doboru naturalnego utrwalającego powolne mutacyjne zmiany. Gdybyśmy w tym przypadku mieli do czynienia z działaniem doboru naturalnego znaczyłoby to, że istnieją mutacje kierunkowe, czego nie dopuszcza ewolucjonizm.
Ponadto co to za ewolucja, w której nie powstał żaden nowy gatunek, czyli to co podobno świadczy o zachodzeniu ewolucji.
Autor cytuje badania, w których 5 par jaszczurek zostało przeniesionych z jednej wyspy, na inna pobliską wysepkę, gdzie wcześniej nie występowały. Po 37 latach, czyli po 18 pokoleniach stwierdzono, że jaszczurki przeniesione nie odżywiają się już owadami, ale są roślinożerne i mają wyraźne przystosowania anatomiczne jelit do trawienia pokarmu roślinnego, które nie występują u populacji wyjściowej. Autor uważa to za dowód ewolucji.
Jasne, że tak. Co prawda to nie powstał żaden nowy gatunek. Po drugie – skąd się wzięły tak szybkie zmiany morfologiczne, tzn., jak duże musiały być zmiany genetyczne i czym miały by być spowodowane, że jaszczurki tak szybko zmieniły fenotyp i stały się wegetarianami? Jedynym wytłumaczeniem – jeśli jest to ewolucja – jest zajście mutacje celowych, których zachodzenie ewolucjonizm wyklucza. Dawkins udowadnia tu coś zupełnie przeciwstawnego temu co chciał udowodnić.
Następnie Dawkins analizuje 'bat na kreacjonistów’ czyli doświadczenia z 12 liniami
Escherichia coli hodowanymi przez 45 tys. pokoleń w laboratorium na podłożu zawierającym agar jako podłoże, cytrynian jako środek konserwujący i glukozę – jako 'jedzonko’ dla bakterii. Bakterie były klonami jednego osobnika.
Należy jeszcze dodać, że dieta była niewystarczająca. Bakterie do rozwoju potrzebują coś więcej niż tylko glukozę, a przede wszystkim sole mineralne, o których autor nic nie wspomina. Warunki nie były więc normalne.
U wszystkich linii przez pierwsze 2 tys. pokoleń obserwowano stopniowy wzrost wielkości osobników – wzrost ten był różny w różnych liniach.
U 2 szczepów (izolowanych), których rozwój przebiegał podobnie, zaobserwowano zmiany genetyczne w ekspresji 59 genów i u obu z nich zmiany te miały identyczny charakter. Nie mogło to być oczywiście sprawą przypadku. Dawkins za badaczami uważa to za 'kumulacyjne działanie doboru naturalnego, który w obu liniach niezależnie – faworyzuje te same zachodzące krok po kroku zmiany’.
Zaobserwowano również że, że jedna z linii po 33 000 pokoleniu nagle zaczęła metabolizować cytrynian, którego normalnie, według Dawkinsa, bakterie nie metabolizują, przynajmniej w obecności tlenu. Badacze wykluczyli możliwość pojedynczej rzadkiej mutacji. Po starannych obserwacjach, stwierdzono, że były to dwie rzadkie, odległe w czasie mutacje, które łącznie razem dały taki efekt. A wiec zaszły dwie przypadkowe mutacje, a następnie zadziałała selekcja. Jednym słowem – ewolucja w czystej postaci.
Gdyby np. kumulacyjne działanie doboru działało na wzrost – zwiększyłby się średni wzrost oraz zmniejszył by się rozstęp między najmniejszym, a największym osobnikiem. Gdyby to było kumulatywne działanie doboru – obserwowano by go we wszystkich szczepach. Niestety nie wiem – jak przejawiała się ekspresja tych genów i co właściwie było identyczne – autor o tym nie pisze – więc nie potrafię powiedzieć nic więcej.
Escherichia coli jest anaerobem fakultatywnym, który w warunkach tlenowych uzyskuje energię przez glikolizę korzystając z tzw. cyklu Krebsa (cykl kwasu cytrynowego), a w warunkach beztlenowych rozkłada glukozę korzystając z cyklu pentozowego.
W procesie glikolizy, glukoza jest przekształcana w kwas pirogronowy, który po przekształceniu w acetylokoenzym A, przechodzi w cytrynian, etc.
Jak widać cytrynian jest normalnie obecny i metabolizowany w komórkach i nie wymaga to jakiejś specjalnej mutacji. Prawdopodobnie kiedy zabrakło glukozy, glikoliza nie mogła się już odbywać więc zerwana została przemiana pirogronianu w acetylokoenzym A, a cały cykl Krebsa był zasilany przez dostawy cytrynianu z zewnątrz. Konieczne było również zapewnienie przepuszczenia cytrynianu przez błonę komórkową. Dwie mutacje z pewnością nie wystarczyłyby aby to spowodować. Jest to wyraźnie zestaw mutacji celowych, dotyczących próby zutylizowania dotychczas nie wykorzystywanego źródła pokarmu. Być może to zaprogramowany mechanizm, który uruchamia się u jednego osobnika na miliard, w przypadku długotrwałego kontaktu z jakimś związkiem. Zmiany, które zaszły u jednego osobnika mogły przenieść się łatwo na inne osobniki, poprzez rekombinację przy rozmnażaniu płciowym albo przez transpozycję.
Dawkins znowu udowadnia coś czemu chciał zaprzeczyć.
W tekście tego rozdziału zawarto jeszcze jedno niepokojące stwierdzenie, że jeżeli mutacje zachodzą losowo np. u jednego osobnika na milion, to przy wielu miliardach osobników uczestniczących w trakcie doświadczeniu musiały zajść wszystkie możliwe mutacje. Z przebiegu i opisu doświadczenia wynika, że nie zaszła nawet część możliwych mutacji, a to przeczy tezie o losowości mutacji. Mutacje – przynajmniej w tym doświadczeniu – wcale nie były losowe.
Gdyby ludzie znaleźli się na stałe w jakimś równie niesprzyjającym środowisku, z pewnością użyli by rozumu i starali się znaleźć sposób na wykorzystanie tak powszechnego czynnika środowiska do własnych celów. Historia ludzka pełna jest takich historii. Bakterie uczyniły to samo, ale użyły zaprogramowanego mechanizmu genetycznego.
Dalej Dawkins opisuje eksperymenty wykazujące ponoć ewolucję zdolności kamuflażu u gupików. Śledzono liczbę i wielkość plamek na ciele osobników w zależności od rodzaju podłoża i różnej presji ze strony drapieżników.
Stwierdzono, że liczba plamek na osobnika, zanim wpuszczono drapieżcę, miał rozkład losowy. W przypadku rozwoju bez drapieżników i umiarkowanej presji ze strony drapieżców liczba plamek rosła, a potem utrzymywała się na stałym wysokim poziomie. Uznano to za przejaw doboru płciowego czyli preferencji samic. W przypadku obecności drapieżców liczba plamek malała.
Wielkość plamek w obecności drapieżników, przy kamienistym dnie – rosła, przy piaszczystym dnie – malała. W nieobecności drapieżców – było odwrotnie. Badania potwierdzono w naturalnych warunkach.
Oczywiście Dawkins nie ma racji. Nie była to żadna ewolucja. W tak krótkim okresie czasu liczonym w latach – nie mogły zajść żadne zmiany genetyczne. Z pewnością nie było to spowodowane mutacjami – ani losowymi ani celowymi. A więc czym? Co takiego zaszło w populacji, że doprowadziło do szybkich zmian wielkości ciała, czy umaszczenia? Jeśli geny się nie zmieniły to co się zmieniło?
Otóż każda populacja poddana działaniu doboru jest w kleszczach dwóch procesów. Z jednej strony zawężenie zmienności, co sprzyja przetrwaniu w stałych warunkach środowiska i rozszerzenie zmienności, które ułatwia przetrwanie, gdy warunki środowiska się zmienią lub osobnik wyemigruje i znajdzie się w innym środowisku. Każdy gatunek zachowuje pewną charakterystyczną dla niego równowagę tych procesów.
Gupiki podobnie jak ptaki bataliony nie mają stałego umaszczenia. W ich umaszczeniu można zaobserwować wielką różnorodność, czyli zmiany barw i umaszczenia zachodzą bardzo łatwo. Można więc przypuszczać, że u gupików istnieją geny regulatorowe, które regulują na bieżąco – nawet w trakcie życia osobnika – jego mimikrę. Może odbywa to się wolniej niż u kameleonów, ale z pewnością jest możliwe. Jeśli nawet dany osobnik nie ma możliwości zmiany umaszczenia to z pewnością jest to możliwe z pokolenia na pokolenie.
Jest zupełnie oczywiste, że wielkość plamek w obecności drapieżców zmieniała się – drapieżniki zjadły silniej wyróżniające się osobniki najpierw. Zjedzony osobnik raczej nie przekaże swych genów dalej.
Jeśli przyjrzeć się wszystkim powyższym przykładom można zauważyć jedną podstawową rzecz – Dawkins pisze o ewolucji, ale nie odróżnia zmian ewolucyjnych od mikroewolucyjnych. Wszystko o czym pisze – to nietrwałe mikroewolucyjne zmiany zachodzące z pokolenia na pokolenia – które są wprowadzane przy zmianach środowiska i mogą być wycofywane – gdy środowisko się zmieni.
Gdyby gatunki, czy osobniki czekały na powolne ewolucyjne mutacyjne zmiany – nie było by na świecie ani jednego gatunku.
Dalej Dawkins wspomina o kilku gatunkach, o których ewolucja zapomniała. Dzisiaj spotykane osobniki niczym nie różnią się od osobników znajdowanych w osadach sprzed setek milionów lat. Dawkins stwierdza, że zwierzęta ewoluują w różnym tempie, a niektóre z nich wcale. Dawkins zaprzecza istnieniu ewolucji. Problem jest następujący. Według ewolucjonistów wszystkie zwierzęta pochodzą od jednego przodka. Mają podobny kod genetyczny, który ulega powolnym zmianom w wyniku mutacji i to generuje zmiany ewolucyjne. Jeżeli kody genetyczne są podobne – to i częstość mutacji powinna być podobna. Nie ma żadnego uzasadnienia dla różnic w szybkości mutacji. Dlaczego Lingula nie ewoluowała, skoro ewoluowali jej bliscy krewni? Morskie środowisko było dla nich niemal identyczne … Środowisko ulegało zmianom, krewni ewoluowali, a u Lingula mutacje nie zachodziły, dobór nie działał, dryfu nie było. Wrogowie ewoluowali, a Lingula nie. Dna morskie zamieniły się w góry – Lingula trwała niezmieniona.
Gdy czyta się o takich gatunkach od razu przychodzi do głowy myśl, którą każdy ewolucjonista odrzuci, a autora myśli uzna od razu za kompletnego idiotę. Zanim to stwierdzisz, przeczytaj książkę do końca. Otóż myśl jest taka: 'Tych gatunków nie było przez miliony lat, a potem pojawiły się ponownie’. 🙂
Brakujące ogniwo? Czemu „brakujące”?
W tym rozdziale Dawkins omawia szereg tematów dotyczących głównie niezrozumienia przez kreacjonistów podstawowych twierdzeń ewolucjonistów. Podstawowym błędem kreacjonistów jest twierdzenie, że ewolucjoniści uważają, że np. małpa wyewoluowała w człowieka, czyli że człowiek pochodzi od małpy. I podobne twierdzenia dotyczące innych gatunków. Podaje również, że znaleziono doskonale zachowany szkielet Darwinius masillae sprzed 47 milionów lat, który jest ogniwem pośrednim między 'lemuropodobnymi’, a 'małpopodobnymi’. To zwierzątko nie jest zbyt podobne do naszego kotozająca, ale nie odbiega od naszych wyobrażeń. Okazuje się, że jednak ogniwa pośrednie istnieją, a drzewko ewolucyjne zaczyna mieć coś więcej niż liście … No i chwała Bogu :-).
To myślenie można rozszerzyć. Jeżeli każdy osobnik różni się genetycznie od rodziców, a zmiany są stopniowe, to na świecie nie ma żadnych gatunków, żadnych ogniw końcowych, a wszystko to co widzimy dookoła nas i wszystkie wykopywane okazy to jedynie stadia pośrednie, które trzeba na schemacie umieścić w odpowiednim miejscu. Osobniki grupujemy w gatunki tylko dla wygody na podstawie stopnia podobieństwa morfologicznego, genetycznego i zdolności krzyżowania.
Inną możliwością jest fakt występowania chorób genetycznych. U człowieka znamy ich ponad 10 000. Być może znajdowane formy to po prostu zwykłe mutanty, a ewolucjoniści uważają je za formy pośrednie.
Dawkins podkreśla tutaj znaczenie obserwacji, że ewolucyjne następstwo gatunków jest zgodne z kolejnością i wiekiem warstw geologicznych, w których są znajdowane i że jak dotąd nie znaleziono żadnego 'szkieletu królika w prekambrze’ co oczywiście obaliłoby teorię ewolucji. Jak na razie wszystkie zwierzątka są na właściwych miejscach.
Wyobraź sobie Czytelniku taką sytuację. Robisz zdjęcia. Niektóre z nich są kiepskie. Kasujesz je. Robisz następne. Znowu część kasujesz. Robisz następne. Nagle okazuje się, że twoja karta jest uszkodzona i nie możesz jej odczytać. Bierzesz program do odzyskiwania danych i nagle okazuje się, że z odzysku otrzymujesz dużo większą liczb zdjęć lub fragmentów zdjęć niż ich ostatnio było. To samo zauważysz odzyskując pliki z twardego dysku. Odzyskasz fragmenty – duże lub małe, plików, o których już dawno zapomniałeś! Miały być zniszczone, ale jednak się zachowały …
Może paleontologia to system odzyskiwania Bożych prototypów, o których pamięć miała zniknąć …
„Brakujące ogniwo” – już nie!
W tym rozdziale Dawkins pisze o hipotetycznej linii ewolucyjnej człowieka. Zasadniczym problemem jest to, że dowody są bardzo niekompletne. Słynna Lucy to tylko parę kości.
Wyobraźmy sobie, że ufoludek z Marsa przylatuje w latającym spodku na Ziemię i robi pomiary i badania różnych osobników ludzkiego gatunku. Trafnie rozpoznaje samce i samice. Ale badając kolory skóry znajduje białych, czarnych, żółtych i czerwonych. Uznaje je za rasy barwne lub podgatunki. Następnie znajduje koszykarza NBA, który poleciał do swoich kumpli Pigmejów na lokalne piwo. Oczywiście zostanie opisany jako inny gatunek. Znajdzie osobniki o wzroście kilkudziesięciu centymetrów jak i o wysokości 2.5 m. Jeżeli znajdziesz w Wikipedii opis hasła 'Julia Pastrana’ zobaczysz osobę o wyraźnych małpich cechach – wynik choroby genetycznej. Została by opisana jako inny gatunek. Podobnie jak wielu biednych osobników chorujących na inne choroby genetyczne, które wywołują fenotypowe skutki. (Oczywiście ciągle są to ludzie, nawet jeśli z różnych względów – genetycznych czy anatomicznych albo estetycznych – nie mogą się krzyżować z innymi osobnikami lub wyglądają odrażająco). Tak właśnie opisują gatunki paleontolodzy. Każda nieco inna czaszka, to nowy gatunek i ogniwo pośrednie.
Gdyby zbadać losowo wybrane czaszki współczesnych osobników – nawet nie wychodząc poza ludzki gatunek – uzyskalibyśmy wiele widocznych linii ewolucyjnych …
Jeśli kiedyś niektórzy dostrzegali kanały na Marsie to zapewne nie ma nic dziwnego w tym, że niektórym zwidują się linie ewolucyjne … Kiedy udoskonalimy narzędzia badawcze okaże się, że to tylko złudzenie.
Tobie zajęło to dziewięć miesięcy!
Autor próbuje opisać tutaj rozwój embrionalny zwierząt i skomplikowany sposób przejścia od jednej komórki do w pełni uformowanego organizmu. Dawkins podkreśla, że wbrew temu co popularnie się twierdzi DNA nie jest ani nie określa bezpośrednio planu ciała i jego późniejszego wyglądu. Twierdzi, że 'instrukcje budowy organizmu same zawierają się w przebiegu tego procesu’. Chodzi oczywiście o proces rozwoju organizmu z pojedynczej komórki.
Następnie dowiadujemy się co nieco o komputerowym symulowaniu niektórych zjawisk biologicznych, np. o naśladowaniu na ekranie zachowania dużego stada lecących szpaków. Nie próbowałem nigdy programować stada szpaków, ale rój owadów – tak. Każdy owad porusza się niezależnie, ale dąży do tego samego celu – jakim jest poruszający się (lub nie) kursor. Sam program jest moim dziełem, napisałem go w Javie, ale sam pomysł roju nie jest mój [64]. Pierwotnie napisany był w języku ActionScript przy wykorzystaniu programu Flash 5. Na ryc. widzimy rój 25 owadów skupiających się wokół kursora, którego nie widać na ryc. Kiedy rój ogląda się w ruchu widać doskonałość symulacji, która do złudzenia przypomina zachowanie prawdziwych owadów rojących się wokół jakiegoś obiektu np. 'g…a’.

ganiających za kursorem
-zrzut z ekranu (rys. Jacek Piechota)
Dalej są rozważania na temat przebiegu embriogenezy, enzymów i strukturze białek oraz o włączaniu i wyłączaniu genów w komórce. Autor nie wyjaśnia skąd dana komórka wie co ma produkować, czyli np. skąd wie czy jest w nerce, czy w trzustce? Dalej są ciekawe informacje o malutkim nicieniu Caenorhabditis elegans – bardzo dokładnie zbadanym organizmie, o którym uczeni wiedzą prawie wszystko. Składa się w postaci larwalnej z 558 komórek i 959 w postaci dorosłej. Prześledzono dokładnie kolejność powstawania i powinowactwo komórek. Autor twierdzi, że za włączanie i wyłączanie genów odpowiada skład chemiczny komórek, dlatego w różnych komórkach włączają się różne geny – i to jest oczywiście nieprawda. Tak może być w razie zmian w środowisku. Środowisko się zmienia, co powoduje zmiany w genach. W komórce jest odwrotnie. Najpierw włączają się geny, a w wyniku tego zmienia się skład chemiczny – co oczywiście może uaktywniać inne geny.
Konkluzja jest taka, że 'rozwojem zarodka rządzą lokalne reguły regulujące interakcje komórek, natomiast rozwojem komórkowym sterują lokalne reguły działające na poziomie cząsteczek i związków chemicznych, zwłaszcza białek’.
Wszystko jest OK., ale to jest wiedza na poziomie lat 60 ubiegłego wieku. Prawdopodobnie wnioski nie są Dawkinsa, tylko autorów badań, których cytuje Dawkins. Autor nic nie mówi o regulacji genetycznej tych procesów. Problem jest o tyle skomplikowany, że geny regulujące są regulowane przez geny regulujące, a te są regulowane przez geny regulujące, wszystkie te geny musiały być zaprogramowane w DNA (oraz wszystkie ich włączenie i wyłączenia, gdyż inaczej żadnej regulacji by nie było. Gdyby nicień miał 10 razy więcej genów niż ma to i tak nie wystarczyłoby informacji do prawidłowego zaprogramowania procesu embriogenezy.
Jak naprawdę przebiega regulacja embriogenezy opowiemy w podrozdziale mówiącym o regulacji genetycznej.
Widać z tego rozdziału, że autor nie jest ekspertem w embriologii, ale to co pisze o programowaniu komputerowym jest jak najbardziej prawdziwe i przekonywujące.
Arka kontynentów
Dalej autor pisze o specjacji. Szczególnie zwraca uwagę na zoogeografię, czyli rozmieszczenie gatunków, a zwłaszcza na występowanie gatunków endemicznych. Autor twierdzi, że gdyby zwierzęta migrowały spod góry Ararat, to powstaje problem, w jaki sposób np. torbacze dostały się do Australii? Nie występują przecież nigdzie indziej?
Istnieje parę możliwych odpowiedzi:
- Ziemia była wtedy jednym kontynentem, a kontynenty rozpłynęły się w różnych kierunkach dopiero po potopie
- Noe lub ktoś z jego rodziny – jakiś czas po potopie – innym statkiem – `porozwoził zwierzęta, tam gdzie było właściwe dla nich miejsce, tylko Biblia o tym nie wspomina
- Noe – jakiś czas po potopie – zbudował mniejsze arki, wsadził na nie zwierzątka i pozwolił im odpłynąć w siną dal o czym też Biblia nie wspomina.
W ok. 130 lat po potopie – 'za dni Pelega’ – rozdzieliła się Ziemia [12] (Rdz 10,25). Nie wiadomo co to znaczy. Być może jakiś proces geologiczny spowodował szybkie rozpłynięcie się kontynentów? Z punktu widzenia współczesnej geologii byłaby to katastrofa na skalę globalną, większa niż sam potop, ale być może nie wiemy jeszcze wszystkiego o geologii, a geologiczne warunki po potopie były bardzo szczególne. Trzeba także dodać że, hebrajskie słowo palag oznacza 'rozdzieliła’.
Drzewo genealogiczne
Autor pisze tu o wszystkim i o niczym, czyli niezbyt konkretnie, o bardzo wielu sprawach. Głownym punktem jest tworzenie drzew genealogicznych na podstawie podobieństwa szkieletów. Dalej autor pisze o homologii i analogii i planach budowy organizmów na przykładzie skorupiaków. Następnie autor pokazuje, że jeśli narysujemy czaszkę np. człowieka, to poddając ją deformacjom (transformacjom) można uzyskać czaski innych kręgowców. 'Świadczące na rzecz ewolucji dowody dostarczane przez badania porównawcze różnych gatunków’ są uznane za 'mocniejsze niż te, które pochodzą z zapisu kopalnego’.
Następnie Dawkins pisze co nieco o tworzeniu drzew filogenetycznych na podstawie podobieństw sekwencji zasad DNA u różnych organizmów, a następnie o ewolucji molekularnej, wspominając również o mutacjach neutralnych.
Moim zdaniem zdaniem
Tworzenie drzew genealogicznych w świecie przyrody jest możliwe, jeśli:
- mutacje genetyczne zachodzą losowo – gdyby mutacje nie zachodziły lub zachodziły nielosowo – nie byłoby ewolucji..
- dobór naturalny utrwala lub usuwa fenotypowe przejawy mutacji
- kod genetyczny jest przekazywany z pokolenia na pokolenie
- istnieją gatunki
- gatunki powstają z innych gatunków dzięki specjacji
Nie ma dokładnej i dobrej definicji gatunku. Jednym słowem: nikt nie wie co to jest gatunek. Specjacja jest procesem powstawania gatunków. Gatunków, czyli właściwie czego?
Głównym kryterium odróżniania gatunków od siebie jest kryterium krzyżowania. Jeżeli osobniki krzyżują się swobodnie – to jest to jeden gatunek. Jeśli nie krzyżują się ze sobą – należą do dwóch różnych gatunków.
Każdy z nas, a szczególnie lekarze, znają nieszczęśliwe ludzkie pary, które nie mogą ze sobą mieć potomstwa (mówimy tu o parach heteroseksualnych). Czy zatem mąż i żona w takim związku należą do różnych gatunków? Oczywiście nie. Tak samo może być w przypadku dowolnych osobników dowolnego gatunku.
Opisano ponad milion gatunków. Wszystkie z nich opisano na podstawie różnic morfologicznych. Opis wygląda tak. Łapiemy żuczka. Mierzymy człony czułków. Np. proporcja I i II członu czułków jest inna niż w standardzie. Opisujemy nowy gatunek. Tak opisują gatunki zoologowie. Trochę inny wzór na pokrywach skrzydeł – to przecież nowy gatunek! Wszystkie te opisy to jedna wielka 'zmyła’.
Gdyby ufoludek z Oriona przyleciał na Ziemię i pobrał próbkę Pigmejów wraz z ich kolegą koszykarzem z NBA, który właśnie wpadł do nich na piwo – opisałby 2 różne gatunki, co oczywiście jest bzdurą mimo wielkich różnic morfologicznych. Gdyby uwzględnił wszystkie choroby o podłożu genetycznym, które wpływają na ludzki fenotyp to staranny ufozoolog wyróżniłby prawie 10 000 gatunków tylko wśród ludzi. Nie licząc naturalnych blondynek :-).
Jeśli nawet 'gatunki’ da się wyróżnić – to nie wiadomo ile z tych gatunków jest naprawdę gatunkami.
Nie znamy żadnego gatunku opisanego na podstawie kryteriów krzyżowania. Jeśli nawet takie gatunki zostały opisane to z pewnością dadzą się policzyć na palcach jednej ręki.
Jedyne gatunki, o których wiemy, że się nie krzyżują to niektóre gatunki zwierząt hodowlanych, które mogą dawać żywotne, ale bezpłodne potomstwo w pierwszym pokoleniu.
Historia zapisana w nas
Autor omawia niektóre narządy ludzkiego ciała i usiłuje udowodnić, że budowa wielu organizmów wskazuje, że jest to raczej 'nieinteligentny projekt’, gdyż myślący projektant zaprojektowałby organizmy inaczej.
Wyścig zbrojeń i „ewolucyjna teodycea”
Jest to rozdział mówiący o ekonomii natury, czyli o wyścigu zbrojeń, czyli o dostosowywaniu się organizmów do – często sprzecznych – wymagań środowiska naturalnego, którego elementem są drapieżce, pasożyty, inne osobniki tego samego gatunku, etc.
„Jest to wniosły zaiste pogląd…”
To rozważania na temat informacji zawartej w przyrodzie.
Podsumowanie

Piśmiennictwo
[12] Chrostowski W (red.). 1998. Biblia w przekładzie księdza Jakuba Wujka z 1599 r., Oficyna Wydawnicza „Vocatio”, Warszawa, 2426 pp.
[44] Dawkins R., 2010. Najwspanialsze widowisko świata. Świadectwa ewolucji. Wydawnictwo CIS, pp. 547
[64] Bhangal S., 2000. Flash 5. ActionScript. Podstawy., Helion, Gliwice, 466pp.
[71] Dixon D i in. 2005. Encyklopedia ewolucji. Ziemia i jej fascynująca historia. Wydawnictwo Debit, Biała Podlaska, 368 pp.
/section>
Czasopismo: Blog Zielonego Smoka
Artykuł: nr 175
Autor: Jacek Piechota